Mire kellenek a nukleinsavak? Nukleinsavak - DNS és RNS
>> Nukleinsavak
1. Mi a sejtmag szerepe a sejtben?
2. Milyen sejtszervecskékhez kapcsolódik az örökletes tulajdonságok átvitele?
3. Milyen anyagokat nevezünk savaknak?
(a latin nucleus - mag szóból) először a leukociták magjában fedezték fel. Ezt követően kiderült, hogy a nukleinsavak minden sejtben megtalálhatók, nem csak a sejtmagban, hanem a citoplazmában és különböző organellumokban is.
Kétféle nukleinsav létezik: dezoxiribonukleinsavak (rövidítve DNS) és ribonukleinsavak (RNS). Az elnevezések különbségét az magyarázza, hogy a DNS-molekula tartalmaz szénhidrát dezoxiribóz, az RNS-molekula pedig ribóz.
A nukleinsavak ms-nukleotidokból álló biopolimerek. A DNS és az RNS nukleotid monomerjei hasonló szerkezetűek.
Minden nukleotid három komponensből áll, amelyeket erős köt össze kémiai kötések. Ez egy nitrogéntartalmú bázis, egy szénhidrát (ribóz vagy dezoxiribóz) és egy maradék foszforsav(9. ábra).


Négy nitrogénbázis létezik: adenin, guanin, citozin vagy timin. Meghatározzák a megfelelő nukleotidok nevét: adenil (A), guanil (G), citidil (C) és timidil (T) (10. ábra).
Mindegyik DNS-szál több tízezer nukleotidból álló polinukleotid.
A DNS-molekula rendelkezik összetett szerkezet. Két spirálisan csavart láncból áll, amelyek teljes hosszukban hidrogénkötésekkel kapcsolódnak egymáshoz. Ezt a DNS-molekulákra jellemző szerkezetet kettős hélixnek nevezik.
Amikor a DNS kettős hélixe kialakul nitrogéntartalmú bázisok az egyik áramkör szigorúan be van helyezve egy bizonyos sorrendben a nitrogénbázisokkal szemben egy másik. Ebben az esetben egy fontos mintázat derül ki: egy másik lánc timinje mindig az egyik lánc adeninjével szemben helyezkedik el, a citozin a guaninnal szemben, és fordítva. Ez azzal magyarázható, hogy az adenin és a timin, valamint a guanin és a citozin nukleotidpárjai szigorúan megfelelnek egymásnak, és kiegészítik, vagy kiegészítik (a latinból, komplementum - addíció) egymást. Az adenin és a timin között mindig kettő, a guanin és a citozin között pedig három hidrogénkötések(11. ábra).
A legkisebb méretű RNS-ek (tRNS-ek) átvitele aminosavakat szállít a fehérjeszintézis helyére.
Az információs vagy mátrix-RNS (i-RNS) a DNS-molekula egyik láncának egy szakaszán szintetizálódik, és a fehérje szerkezetére vonatkozó információkat továbbítja a sejtmagból a riboszómákba, ahol ez az információ megvalósul.
És így, Különféle típusok Az RNS-ek egyetlen funkcionális rendszer megvalósítására irányul örökletes információk fehérjeszintézis révén.
Az RNS-molekulák a sejt magjában, citoplazmájában, riboszómáiban, mitokondriumaiban és plasztidjaiban találhatók.
Nukleinsav. Dezoxiribonukleinsav vagy DNS. Ribonukleinsav, vagy RNS, Nitrogénbázisok: adenin, guanin, citozin, timin, uracil. Komplementaritás. Transzfer RNS (tRNS). Riboszomális RNS(rRNS). Messenger RNS (mRNS). Nukleotid. Kettős spirál.
1. Milyen a nukleotid szerkezete?
2. Milyen a DNS-molekula szerkezete?
3. Mi a komplementaritás elve?
4. Milyen hasonlóságok és különbségek vannak a DNS- és RNS-molekulák szerkezetében?
5. Milyen típusú RNS-molekulákat ismer? Mik a funkcióik?
Kamensky A. A., Kriksunov E. V., Pasechnik V. V. Biológia 9. osztály
A honlap olvasói küldték be
Egy élő szervezetben három fő makromolekula található: fehérjék és kétféle nukleinsav. Nekik köszönhetően az egész szervezet létfontosságú tevékenysége és megfelelő működése megmarad. Mik azok a nukleinsavak? Miért van rájuk szükség? Erről bővebben a cikk későbbi részében.
Általános információ
A nukleinsav egy biopolimer, nagy molekulájú szerves vegyület, amelyet nukleotidmaradékok képeznek. Nemzedékről nemzedékre való átvitel genetikai információ - a fő feladat amelyet nukleinsavak hajtanak végre. Az alábbi bemutatóból kiderül ezt a koncepciót részletekben.
A tanulmány története
Az első vizsgált nukleotidot szarvasmarha izomzatából izolálták 1847-ben, és „inozinsavnak” nevezték el. A kémiai szerkezet tanulmányozása során kiderült, hogy ez egy ribozid-5′-foszfát, és N-glikozidos kötést tartalmaz 1868-ban egy „nuklein” nevű anyagot fedeztek fel. Friedrich Miescher svájci kémikus fedezte fel bizonyos biológiai anyagok kutatása során. Ez az anyag foszfort tartalmazott. A kapcsolat megvolt savas tulajdonságokés nem bomlik le proteolitikus enzimek hatására.

Az anyag a C29H49N9O22P3 képletet kapta. A nukleinnek az örökletes információ továbbításában való részvételére vonatkozó feltételezést annak eredményeként terjesztették elő, hogy felfedezték kémiai összetételének a kromatinnal való hasonlóságát. Ez az elem a kromoszómák fő összetevője A „nukleinsav” kifejezést először Richard Altmann vezette be 1889-ben. Ő volt az, aki a vizsgálat során elkészítette azt a módszert, amellyel ezeket az anyagokat fehérje szennyeződések nélkül állítják elő lúgos hidrolízis nukleinsavakat, Levin és Jacob azonosította a folyamat termékeinek fő összetevőit. Kiderült, hogy nukleotidok és nukleozidok. 1921-ben Lewin felvetette, hogy a DNS-nek tetranukleotid szerkezete van. Ez a hipotézis azonban nem igazolódott be, és tévesnek bizonyult.

Osztályozás
A nukleinsavak két típusból állnak: DNS és RNS. Jelenlétük minden élő szervezet sejtjében megtalálható. A DNS főként a sejtmagban található. Az RNS a citoplazmában található. 1935-ben a DNS lágy fragmentációja során 4 DNS-képző nukleotidot kaptak. Ezek a komponensek kristályos állapotban vannak bemutatva. 1953-ban Watstone és Crick megállapította, hogy a DNS igen kettős spirál.
Kiválasztási módszerek

Fejlett különböző módokon vegyületek kinyerése innen természetes források. Ezeknek a módszereknek a fő feltételei a nukleinsavak és fehérjék hatékony szétválasztása, a folyamat során nyert anyagok legkisebb fragmentációja. Ma a klasszikus módszert széles körben alkalmazzák. Ennek a módszernek a lényege a biológiai anyag falainak tönkretétele és további anionos tisztítószerrel történő kezelése. Az eredmény fehérjecsapadék, miközben a nukleinsavak oldatban maradnak. Egy másik módszert is alkalmaznak. Ebben az esetben a nukleinsavakat etanol és sóoldat felhasználásával gél állapotba lehet kicsapni. Ennek során bizonyos óvatossággal kell eljárni. Különösen az etanolt kell nagy körültekintéssel hozzáadni a sóoldathoz, hogy gélcsapadékot kapjunk. Azt, hogy a nukleinsav milyen koncentrációban szabadul fel, milyen szennyeződések vannak benne, spektrofotometriás módszerrel határozható meg. A nukleinsavakat a nukleáz könnyen lebontja, ami az speciális osztály enzimek. Egy ilyen kiválasztásnál szükséges, hogy laboratóriumi felszerelés kötelező gátló kezelésen esett át. Ezek közé tartozik például egy DEPC-inhibitor, amelyet RNS izolálására használnak.
Fizikai tulajdonságok
A nukleinsavak jól oldódnak vízben, de szinte oldhatatlanok szerves vegyületekben. Ezenkívül különösen érzékenyek a hőmérsékletre és a pH-szintre. Nukleinsav molekulák magas molekuláris tömeg, hatására a nukleáz széttöredezhet mechanikai erők. Ezek közé tartozik az oldat összekeverése és rázása.
Nukleinsavak. Felépítés és funkciók

A kérdéses vegyületek polimer és monomer formái megtalálhatók a sejtekben. A polimer formákat polinukleotidoknak nevezzük. Ebben a formában a nukleotidláncokat foszforsav-maradék köti össze. A kétféle heterociklusos molekula, az úgynevezett ribóz és dezoxiribóz tartalma miatt a savak ribonukleinsavak, illetve dezoxiribonukleinsavak. Segítségükkel megtörténik az örökletes információk tárolása, továbbítása és megvalósítása. A nukleinsavak monomer formái közül a legnépszerűbb az adenozin-trifoszforsav. Részt vesz a jelátvitelben és a sejt energiatartalékainak biztosításában.
DNS
A dezoxiribonukleinsav egy makromolekula. Segítségével megtörténik a genetikai információ átvitelének és megvalósításának folyamata. Ezek az információk az élő szervezet fejlődéséhez és működéséhez szükségesek. Az állatokban, növényekben és gombákban a DNS a sejtmagban található kromoszómák része, és megtalálható a mitokondriumokban és a plasztidokban is. Baktériumokban és archaeákban a dezoxiribonukleinsav molekula belülről tapad a sejtmembránhoz. Az ilyen szervezetekben főként cirkuláris DNS-molekulák vannak jelen. Ezeket "plazmidoknak" nevezik. Kémiai szerkezete szerint a dezoxiribonukleinsav nukleotidokból álló polimer molekula. Ezek az összetevők viszont nitrogénbázist, cukrot és foszfátcsoportot tartalmaznak. Az utolsó két elemnek köszönhető, hogy a nukleotidok között kötés jön létre, láncokat hozva létre. Alapvetően a DNS-makromolekula két láncból álló spirál formájában jelenik meg.

RNS
A ribonukleinsav egy hosszú lánc, amely nukleotidokból áll. Nitrogénbázist, ribózcukrot és foszfátcsoportot tartalmaznak. A genetikai információt nukleotidszekvencia segítségével kódolják. Az RNS-t a fehérjeszintézis programozására használják. A ribonukleinsav a transzkripció során keletkezik. Ez az RNS-szintézis folyamata egy DNS-templáton. Speciális enzimek részvételével fordul elő. Ezeket RNS polimerázoknak nevezik. Ezt követően a templát ribonukleinsavak részt vesznek a transzlációs folyamatban. Az RNS-mátrixon így megy végbe a fehérjeszintézis. Aktív részvétel A riboszómák részt vesznek ebben a folyamatban. A fennmaradó RNS-ek kémiai átalakuláson mennek keresztül a transzkripció befejezéséhez. A bekövetkező változások eredményeként a ribonukleinsav másodlagos és harmadlagos szerkezete alakul ki. Az RNS típusától függően működnek.
Az öröklődés és változékonyság molekuláris alapja
1. Nukleinsavak, szerkezetük, funkcióik és keletkezésük
2. A fehérje bioszintézis főbb szakaszai. A genetikai kód alapvető tulajdonságait
3. A génexpresszió szabályozása
Nukleinsavak, szerkezetük és funkcióik
Nukleinsavak lineáris, el nem ágazó heteropolimerek, amelyek monomerjei nukleotidokösszefüggő foszfodiészter kötések.
Nukleotidok- Ezt szerves anyag, melynek molekulái egy pentóz-maradékból (ribóz vagy dezoxiribóz) állnak, amelyhez kovalensen kötődik egy foszforsav-maradék és egy nitrogéntartalmú bázis. A nukleotidokban lévő nitrogénbázisokat két csoportra osztják: purin(adenin és guanin) és pirimidin(citozin, timin és uracil). Dezoxiribonukleotidok tartalmazza dezoxiribóz adenin(A), guanin(G), timin(T), citozin(C). Ribonukleotidok tartalmazza ribózés az egyik nitrogéntartalmú bázis: adenin(A), guanin(G), uracil(U), citozin(C).
Egyes esetekben a felsorolt nitrogénbázisok különféle származékai találhatók a sejtekben - kisebb bázisok, amelyek kisebb nukleotidok részét képezik.
A szabad nukleotidok és a hozzájuk hasonló anyagok játszanak fontos szerep az anyagcserében. Például a NAD (nikotinamid-adenin-dinukleotid) és a NADP (nikotinamid-adenin-dinukleotid-foszfát) elektron- és protonhordozóként szolgál.
A szabad nukleotidok 1...2 további foszforcsoport hozzáadására, kialakítására képesek nagy energiájú vegyületek. A sejt univerzális energiaforrása az ATP - adenozin-trifoszforsav, amely adeninből, ribózból és három foszforsav (pirofoszforsav) maradékból áll. Egy terminális pirofoszfát kötés hidrolízise során körülbelül 30,6 kJ/mol (vagy 8,4 kcal/mol) szabad energia szabadul fel, amelyet a sejt felhasználhat. Ezt a pirofoszfát kötést ún makroergikus(nagy energia).
Az ATP-n kívül más, nukleotid alapú, nagy energiájú vegyületek is léteznek: GTP (guanint tartalmaz; részt vesz a fehérjék és glükóz bioszintézisében), UTP (uracilt tartalmaz; részt vesz a poliszacharidok szintézisében).
A nukleotidok ciklikus formák kialakítására képesek, például cAMP, cCMP, cGMP. A ciklikus nukleotidok különféle élettani folyamatok szabályozóiként működnek.
Nukleinsavak
Kétféle nukleinsav létezik: DNS ( Dezoxiribonukleinsav) és RNS ( ribonukleinsav). A nukleinsavak biztosítják a genetikai (örökletes) információk tárolását, reprodukálását és megvalósítását. Ez az információ nukleotid szekvenciák formájában tükröződik (kódolva). A nukleotidszekvencia különösen a fehérjék elsődleges szerkezetét tükrözi (lásd alább). Az aminosavak és az azokat kódoló nukleotidszekvenciák közötti megfelelést ún genetikai kód . Mértékegység genetikai kód A DNS és az RNS az hármas– három nukleotidból álló szekvencia.
A nukleinsavak kémiailag aktív anyagok. Különféle vegyületeket képeznek a fehérjékkel - nukleoproteinek, vagy nukleoproteinek.
Dezoxiribonukleinsav (DNS) egy nukleinsav, amelynek monomerjei dezoxiribonukleotidok. A DNS az örökletes információ elsődleges hordozója. Ez azt jelenti, hogy minden információ az egyes sejtek felépítéséről, működéséről és fejlődéséről, ill egész szervezet DNS nukleotid szekvenciák formájában írva.
A nukleinsavakat Miescher fedezte fel 1868-ban. Feulgen azonban csak 1924-ben bizonyította be, hogy a DNS a kromoszómák lényeges összetevője. 1944-ben Avery, McLeod és McCarthy megállapította, hogy a DNS kulcsfontosságú szerepet játszik az örökletes információk tárolásában, továbbításában és megvalósításában.
A DNS-nek többféle típusa létezik: A, B, Z, T formák. Ezek közül a B-forma általában a sejtekben található - egy kettős jobboldali hélix, amely két szálból (vagy láncból) áll, amelyeket hidrogénkötések kapcsolnak össze. Mindegyik szálat váltakozó dezoxiribóz és foszforsav csoportok képviselik, a dezoxiribózhoz kovalensen kötődő nitrogénbázissal. Ebben az esetben a két DNS-szál nitrogénbázisai egymás felé irányulnak, és a hidrogénkötések kialakulása miatt alakulnak ki. komplementer párok: A=T (két hidrogénkötés) és G≡C (három hidrogénkötés). Ezért ezeknek a láncoknak a nukleotid szekvenciái egyértelműen megfelelnek egymásnak. A kettős hélix fordulat hossza 3,4 nm, a szomszédos nitrogénbázispárok távolsága 0,34 nm, a kettős hélix átmérője 1,8 nm.
Az eukarióta sejtekben a DNS nukleoprotein komplexek formájában létezik, amelyek hisztonfehérjéket tartalmaznak.
A DNS hosszát a nukleotidpárok számával mérjük (rövidítve - Hétfő, vagy b). Egy DNS-molekula hossza több ezer között mozog Hétfő(rövidítve - TPN, vagy Kb) akár több millió Hétfő (MPN, vagy Mb).
Friedrich Miescher több mint száz évvel ezelőtt (1869-ben) gennysejtek tanulmányozása közben izolált e sejtek magjából. új típusú kémiai vegyületek, amelyet összefoglalóan "nukleinnek" nevezett. Ezek az anyagok, amelyeket később nukleinsavaknak neveztek, savas tulajdonságokkal rendelkeztek, szokatlanul gazdagok foszforban, valamint szenet, oxigént, hidrogént és nitrogént is tartalmaztak. Későbbi vizsgálatuk kimutatta, hogy kétféle nukleinsav létezik: dezoxiribonukleinsav (DNS) és ribonukleinsav (RNS), amelyek szerves részeösszetett fehérjék - az állatok, baktériumok, vírusok, növények összes sejtjében található nukleoproteinek.
A nukleoproteinek [illetve a dezoxiribonukleoproteinek (DNP) és ribonukleoproteinek (RNP-k)] összetételükben, méretükben és fizikai-kémiai tulajdonságaikban különböznek egymástól. A nukleoproteinek nevei csak a nukleinsavak részét képező szénhidrát komponens (pentóz) természetét tükrözik. Az RNP-ben a szénhidrátot ribóz, a DNP-ben dezoxiribóz képviseli. A "nukleoproteinek" elnevezés a sejtmag nevéhez kapcsolódik, ahol először fedezték fel őket. Mostanra azonban megállapították, hogy a DNP és az RNP más szubcelluláris struktúrákban is megtalálható. Ebben az esetben a DNP-k túlnyomórészt a sejtmagban, az RNP-k pedig a citoplazmában lokalizálódnak. Ugyanakkor a DNP-k nyitottak a mitokondriumokban, és a nagy molekulatömegű RNP-k a sejtmagokban és a sejtmagokban is megtalálhatók.
| A DNS és az RNS közötti különbségek | ||
| Mutatók | DNS | RNS |
| Elhelyezkedés | sejtmag, a kromatin része, kevés a mitokondriumban (a teljes DNS 0,2%-a) | minden részében |
| Cukor (pentóz) | Dezoxiribóz | Ribóz |
| Nitrogéntartalmú bázisok | Adenin, guanin, citozin, Timin |
Adenin, guanin, citozin, Uracil |
| A láncok száma egy molekulában | 99,99% - kettős hélix, 0,01% egyszálú | 99,99% - egyszálú, 0,01% kétszálú |
| Molekula alakja | Minden egyszálú gyűrű alakú. A legtöbb kétszálú lineáris, néhány gyűrűs. |
Lineáris molekulák |
Nukleinsavak kémiai összetétele
A nukleinsavak izolálása a fehérjékkel alkotott komplexükből, majd az azt követő teljes hidrolízisük lehetővé tette a kémiai összetétel nukleinsavak. Így a teljes hidrolízis során a hidrolizátumban purin és pirimidin bázisok, szénhidrátok (ribóz és dezoxiribóz) és foszforsavat találtak.
Nitrogénbázisok (N-bázisok)
A purin és pirimidin bázisok szerkezete két aromás bázison alapul heterociklusos vegyületek- purin és pirimidin. A perimidin molekula egy heterociklust tartalmaz. A purin molekula két fuzionált gyűrűből áll: pirimidinből és imidazolból.
Figyelj! A nitrogéntartalmú bázisok aromás gyűrűjében az atomok számozása arab számokkal történik prímszám nélkül [ " ]. A [ " ] szimbólum (ejtsd: "prím" vagy "prim") azt jelzi, hogy a megfelelő szám a pentóz atomjait jelöli gyűrű, például 1" (lásd alább).

A nukleinsavak összetételében három fő pirimidinbázis található: citozin (C), uracil (U) és timin (T):

és két purin - adenin (A) és guanin (G)

Az egyik fontos tulajdonságait A nitrogéntartalmú bázisok (hidroxicsoportokat tartalmaznak) két tautomer formában való létezésének lehetősége, különösen laktim és laktám formában, a tápközeg pH-értékétől függően. A tautomer átalakulások az uracil példájával ábrázolhatók.

Kiderült, hogy a nukleinsavak összetételében a purinok és pirimidinek összes hidroxi-származéka laktám formában van.
A főbb bázisokon kívül ritka (kisebb) nitrogénbázisokat fedeztek fel a nukleinsavakban. Kisebb bázisok túlnyomórészt itt találhatók transzport RNS-ek, ahol listájuk megközelíti az 50-et, nyomokban riboszomális RNS-ben és DNS-ben. A transzfer RNS-ekben a kisebb bázisok az összes nukleotid 10%-át teszik ki, aminek nyilvánvalóan fontos fiziológiai jelentése van (az RNS-molekula megvédése a hidrolitikus enzimek hatásától). A kisebb bázisok közé tartoznak továbbá a metilezett purin- és pirimidinbázisok, például 2-metil-adenin, 1-metil-guanin, 5-metil-citozin, 5-hidroxi-metil-citozin stb.


Szénhidrát
A nukleinsavakban található szénhidrátok (pentózok) a ribóz és a 2-dezoxiribóz, amelyek β-D-ribofuranóz formában vannak (képletek a bal oldalon).
Néhány fág DNS-ben glükózmolekulát találtak, amely glikozidos kötéssel kapcsolódik az 5-hidroxi-metil-citozinhoz.
A szénhidrát ciklus konformációja (pentózok)
A nukleinsavak szénhidrátciklusa (pentóz) számára energetikailag kedvezőtlen a síkbeli konformáció, amikor a C1, C2, C3, C4 szénatomok és az oxigénheteroatom egy síkban vannak. Ezen aminosavak számos elméletileg lehetséges konformációja közül csak kettő valósul meg a polinukleotidokban: vagy a C2"-endokonformáció, vagy a C3"-endokonformáció. Ezek a konformációk a C4" kötés körüli forgásból származnak, ami a gyűrű olyan torzulásához vezet, amelyben a pentóz atomok egyike (egy öttagú furanózgyűrű) a másik négy atom által létrehozott síkon kívül jelenik meg. Ez a konformáció endo- vagy exo-struktúra, attól függően, hogy lokalizálódik adott atom a sík ugyanazon az oldalán, mint a C5", vagy az ellenkező oldalon.
Azokat az anyagokat, amelyekben nitrogéntartalmú bázisok pentózzal kombinálódnak, nukleozidoknak nevezzük (2. ábra).

A nukleozidokat N-glikozidok közé sorolják. Pirimidin nitrogéntartalmú bázisaik (egy heterociklus) glikozidos kötéssel kapcsolódnak a pentózhoz N-1-en, purinbázisaik pedig N-9-en keresztül. A pentóz típusától függően kétféle nukleozidot különböztetnek meg - a 2-dezoxiribózt tartalmazó dezoxiribonukleozidokat és a ribózt tartalmazó ribonukleozidokat.
A dezoxiribonukleozidok csak a DNS-ben, a ribonukleozidok pedig csak az RNS-ben találhatók. A pirimidin és purin nukleozidok megfelelő nitrogénbázisokat tartalmaznak:

A főbbeken kívül vannak kisebb nukleozidok, amelyek kisebb nitrogénbázisokat tartalmaznak. A legtöbb kisebb nukleozidot a tRNS tartalmazza. Az összes tRNS-ben található leggyakoribb kisebb nukleozidok a dihidrouridin, a pszeudouridin (rövidítve Ψ) és a ribotimidin. A pszeudouridinből hiányzik a szokásos N-glikozid kötés. Ebben a ribóz C-1 atomja kapcsolódik az uracil C-5 atomjához.
Sztérikus okok miatt a purin bázisok a DNS-ben található purin nukleotidokban csak két, sztérikusan hozzáférhető konformációt tudnak felvenni a dezoxiribóz-maradékhoz képest, amelyeket szin konformációnak és antikonformációnak neveznek.
Ugyanakkor a pirimidin nukleotidok pirimidinbázisai antikonformerek formájában vannak jelen a DNS-ben, ami a nukleotid szénhidrát része és a karbonil-oxigén között fellépő sztérikus inkonzisztenciákhoz kapcsolódik a C-2 pozícióban. pirimidin. Emiatt a pirimidinbázisok főként antikonformációt kapnak (Nelson D.L., Cox M.M., Lehninger Principles of Biochemistry, W.H. Freeman (szerk.), San Francisco, 2004).
A nukleotidok a megfelelő típusú nukleozidok foszforsavval alkotott vegyületei. Ribózt tartalmazó ribonukleotidokra és 2-dezoxiribózt tartalmazó dezoxiribonukleotidokra is fel vannak osztva. A nukleotid neve a nitrogéntartalmú bázis típusából és a foszforsavmaradékok számából származik. Ha egy foszforsav-maradékot tartalmaz - nukleozid-monofoszfátot (például dAMP-dezoxiadenozin-monofoszfátot), két maradékot - nukleozid-difoszfátot (például dADP-dezoxiadenozin-difoszfátot), három maradékot - nukleozid-trifoszfátot (például dezoxiadenozin-AT-trifoszfátot). A dezoxiribóz 5"-os szénatomjához foszforsavat adnak, és α, β, γ jelöléssel látják el.
Az alábbiakban az adenil-nukleotidok szerkezete látható.
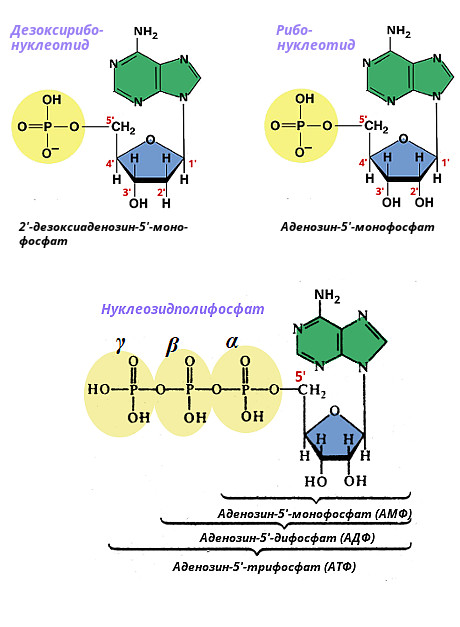
Foszfát adható hozzá különböző pozíciókat pentózgyűrűk (ribonukleotidokban - a 2", 3", 5", dezoxiribonukleotidokban - a 3", 5" pozíciókban). A sejtben jelen lévő szabad nukleotidok foszfátcsoportot tartalmaznak az 5" pozícióban. A nukleozid 5"-foszfátok részt vesznek a nukleinsavak biológiai szintézisében, és lebontásuk során keletkeznek. Mivel a nukleozid 5"-foszfátok vagy mononukleotidok a megfelelő nukleozidok származékai, ugyanazokat a fő és ritka ribomononukleotidokat és dezoxiribomononukleotidokat különböztetjük meg.
A mononukleotid foszfátvégének további foszfátok hozzáadásával történő megnyúlása nukleozid-polifoszfátok képződéséhez vezet. A sejtekben leggyakrabban nukleozid-difoszfátok és nukleozid-trifoszfátok találhatók. A nukleozid-foszfátok nevei és rövidítései az alábbiakban találhatók:
Az összes nukleozid-foszfát anionok formájában található a sejtben, ezért az adenozin-foszfátokat pontosabban AMP 2-, ADP 3-, ATP 4- néven nevezzük. Az ADP és az ATP makroerg, azaz energiában gazdag vegyületek, amelyek kémiai energiáját a szervezet különféle funkciókra használja fel. A fennmaradó nukleozid-di- és trifoszfátok szintén részt vesznek a biológiai anyagok szintézisének reakcióiban.
Nemzetközi szabványos rövidítések
A nukleinsavak vizsgálatával foglalkozó munkákban az Általános és Alkalmazott Kémia Nemzetközi Szövetsége (IUPAC) és a Biokémikusok Nemzetközi Szövetsége (IUB) bizottsága által javasolt atomszámozási sémákat és rövidítéseket alkalmazzák. Az IUPAC-IUB albizottság közös szabvány definíciók(IUPAC-IUB, 1983).
A bázisok, nukleozidok és nukleotidok jelölésére használt rövidítések és szimbólumok (Arnott S., 1970).
| Bázis | |||||
| Név | Szimbólum | Név | Szimbólum | Név | Szimbólum |
| 1. Ribonukleozidok és ribonukleotidok | |||||
| Uracil | Ura | Uridine | Urd vagy U | Uridilsav | 5"-UMP vagy pU |
| Citozin | Cyt | Cytidin | Cyd vagy C | Cytidilsav | 5"-CMP vagy PC |
| Adenin | Ade | Adenozin | Ado vagy A | Adenilsav | 5"-AMP vagy pA |
| Guanin | Gua | Guanozin | Guo vagy G | Guanilsav | 5"-GMP vagy pG |
| 2. Dezoxiribonukleozidok és dezoxiribonukleotidok | |||||
| Timin | Tiéd | Dezoxitimidin | dThd vagy dT | Dezoxitimidilsav | 5"-dTMP vagy pdT |
| Citozin | Cyt | Dezoxicitidin | dCyd vagy dC | Dezoxicitidinsav | 5"-dCMP vagy pdC |
| Adenin | Ade | Dezoxiadenozin | dAdo vagy dA | Dezoxiadenilsav | 5" dAMP vagy pdA |
| Guanin | Gua | Dezoxiguanozin | dGuo vagy dG | Dezoxiguanilsav | 5" dGMP vagy pdG |
| 3. Polinukleotidok | |||||
Az azonos típusú nukleotidokból álló szintetikus polimereket homopolimereknek nevezzük. Megnevezés például poliadenilsav - poli(A) A váltakozó nukleotidszekvenciájú szintetikus polimereket heteropolimereknek nevezzük. A váltakozó dA-t és dT-t tartalmazó kopolimert - poli(dezoxiadenilát-dezoxitimidilát) poli d(A-T) vagy poli(dA-dT) vagy (dA-dT) vagy d(A-T)n jelöléssel látjuk el. A dA, dT véletlenszerű kopolimernél kötőjel helyett vesszőt kell a karakterek közé tenni, például poli d(A,T). A komplementer duplex kialakulását egy pont jelzi a szimbólumok között - poly(dA) · poli(dT); hármas hélix - poli(dA) · 2poly(dT). Az oligonukleotidokat jelöljük a következő módon: például a guanilil-3",5"-citidil-3",5"-uridin - GpCpU vagy GCU oligonukleotid, ahol az 5"-terminális nukleotid G, a 3"-terminális nukleotid pedig U. A komplementer kapcsolt oligonukleotidok nómenklatúrája a következő: |
|||||

Az 5. ábrán. Bemutatjuk a nukleotidokhoz alkalmazott atomszámozási rendszert. A cukoratomokat jelző szimbólumokat a bázisatomok szimbólumaitól egy "elsődleges" szimbólum különbözteti meg. A polinukleotid gerincet a P -> O5" -> C5" -> C4" -> C3" -> O3" -> P irányban írjuk le.
A cukorgyűrűben a számozás a következő: C1" -> C2" -> C3" -> C4" -> O4" -> C5".
A dezoxiribózban a C5" atomnál és a C2" atomnál lévő két hidrogénatom, valamint a foszforatomoknál a két szabad oxigénatom 1-es és 2-es számmal van rendelve, és ez a következőképpen történik: ha végignézünk a láncon az O5"-> C5" irányba, majd az óramutató járásával megegyező irányba haladva egymás után haladunk át a C4", H5"1, H5"2 atomokon. Hasonlóan, ha a lánc mentén O3" -> P - O5 irányba nézünk ", akkor az óramutató járásával megegyező irányban haladva egymás után áthaladunk az O5 atomokon", Op1, Op2.
A nukleinsavak általános jellemzői
A nukleinsavak vagy polinukleotidok olyan nagy molekulatömegű anyagok, amelyek 3", 5"-os foszfodiészter kötésekkel láncba kapcsolt mononukleotidokból állnak..
A sejtek teljes DNS- és RNS-tartalma attól függ funkcionális állapot. A spermiumban a DNS mennyisége eléri a 60%-ot (száraz sejttömegre vonatkoztatva), a legtöbb sejtben 1-10, az izmokban pedig körülbelül 0,2%. Az RNS-tartalom általában 5-10-szer nagyobb, mint a DNS. Az RNS/DNS arány a májban, a hasnyálmirigyben, az embrionális szövetekben és általában az aktív fehérjét szintetizáló szövetekben 4 és 10 között van. A mérsékelt fehérjeszintézissel rendelkező szövetekben az arány 0,3 és 2,5 között mozog. A vírusok különleges helyet foglalnak el. Genetikai anyaguk lehet DNS (DNS vírusok) vagy RNS (RNS vírusok).
Azokban a baktériumsejtekben, amelyek nem rendelkeznek maggal (prokarióták), a DNS-molekula (kromoszóma) a citoplazma egy speciális zónájában - a nukleoidban - található. Ha a baktérium sejtmembránjához kapcsolódik, akkor mezoszómának nevezik. Egy kisebb DNS-fragmens ezen a kromoszómális zónán kívül található. A baktériumokban lévő DNS ilyen szakaszait plazmidoknak vagy episzómáknak nevezzük. A sejtmaggal (eukarióták) rendelkező sejtekben a DNS megoszlik a mag között, ahol a kromoszómák és a sejtmag része, valamint az extranukleáris organellumok (mitokondriumok és kloroplasztok) között. Vannak megfigyelések, amelyek szerint nagyon kis mennyiségű DNS van jelen a mikroszómákban.
A sejt DNS-ének körülbelül 1-3%-a extranukleáris DNS, a többi pedig a sejtmagban koncentrálódik. Ez azt jelenti, hogy az örökletes tulajdonságok nemcsak a sejtmagra, hanem a sejtek mitokondriumaira és kloroplasztiszaira is jellemzőek. Az érett tojásokat szokatlanul magas extranukleáris DNS-tartalom jellemzi, amelyben számos mitokondriumban és tojássárgája lemezben jelen van, az utóbbiakban pedig nem. genetikai anyag, hanem a nukleotidok tartaléka.
Az RNS a DNS-sel ellentétben egyenletesebben oszlik el a sejtben. Ez a körülmény önmagában arra utal, hogy az RNS funkciója dinamikusabb és változatosabb. A sejtekben magasabb rendű szervezetek az összes RNS körülbelül 11%-a a sejtmagban, körülbelül 15%-a a mitokondriumokban, 50%-a a riboszómákban és 24%-a a hialoplazmában található.
A DNS molekulatömege az élő objektum összetettségi fokától függ: baktériumokban 2 10 9, emberekben és állatokban eléri a 10 11-et. A baktériumokban a DNS egyetlen óriási molekula formájában található, amely lazán kötődik a fehérjékhez. Más objektumokban a DNS-t fehérjék vagy egyszerű aminok veszik körül. A vírusokban ezek a legegyszerűbb bázikus fehérjék vagy poliaminok (putreszcin és spermidin), amelyek semlegesítik negatív töltés A DNS-molekulák foszfátcsoportjaihoz kötődve. Egyes állatok és halak spermájában a DNS komplexeket képez protaminokkal és hisztonszerű fehérjékkel. Az emberi sejtek és más magasabb rendű szervezetek kromoszómáiban a DNS hisztonokhoz és nem hiszton fehérjékhez kapcsolódik. Az ilyen fehérje-DNS komplexeket dezoxiribonukleoproteineknek (DNP) nevezik.
Az RNS molekulatömege lényegesen kisebb, mint a DNS. Az elvégzett funkciótól, a molekulatömegtől és a nukleotidok összetételétől függően az RNS következő fő típusait különböztetjük meg: információs vagy templát (mRNS), transzport (tRNS) és riboszomális (rRNS). A különböző rRNS-ek molekulatömege különbözik (13. táblázat). A három fő típuson kívül vannak kisebb, vagy ritka RNS-ek, amelyek sejttartalma elenyésző, funkciójukat még csak tanulmányozzák.
A legtöbb RNS-típus a sejtben a különféle fehérjék. Az ilyen komplexeket ribonukleoproteineknek (RNP) nevezik. A nukleinsavak jellemzőit a táblázat foglalja össze. 1.
| Asztal 1. rövid leírása magasabb rendű szervezetek sejtjeinek nukleinsavai | |||||
| Nukleinsav típus | Molekulatömeg | Ülepítési állandó (Svedberg-S egységekben) | Tartalom a cellában, % | Lokalizáció a cellában | Funkció |
| DNS | 10 11 | - | Az összes DNS 97-99%-a A teljes DNS 1-3%-a | Mag Mitokondriumok | Genetikai információ tárolása és részvétel a szülői DNS átvitelében a sejtosztódás során vagy az RNS átvitelében az életfolyamatok során |
| mRNS | 4 10 4 - 1,2 10 6 | 6-25 | A teljes RNS 25%-a | Mag, citoplazma | Ez egy olyan DNS-szakasz másolata, amely információkat tartalmaz egy fehérje polipeptidláncának szerkezetéről. Információt visz át a DNS-ből a fehérjeszintézis helyére - a riboszómákba |
| tRNS | 2,5 10 4 | ~4 | A teljes RNS 15%-a | Hialoplazma, riboszómák, mitokondriumok | Részt vesz az aminosavak aktiválásában, a riboszómákba történő szállításában és a polipeptidek összeállításában aminosavakból a riboszómákon |
| rRNS | 0,7 10 6 | 18 | Az összes RNS 80%-a | A citoplazma riboszómái | A citoplazmában (vagy mitokondriumban) riboszómák vázát képezi, amelyet riboszómális fehérjék borítanak be. Segédszerepet játszik a fehérjék riboszómákon történő összeállításában |
| 0,6 10 6 | 16 | A mitokondriumok riboszómái | |||
| ~4 10 4 | 5 | Minden riboszóma | |||
| Kromoszóma vektor RNS | 10 4 | 3 | Lábnyomok | Nukleáris kromoszómák | DNS gének felismerése és aktiválása |
| Alacsony molekulatömegű nukleáris RNS-ek | 2,5 10 4 -5 10 4 | 4-8 | A százalék töredékei | A citoplazma sejtmagjai, RNP részecskéi | DNS gének aktiválása, a tRNS-t a sejtmagból a citoplazmába szállító fehérjerészecskék vázának kialakulása |
A nukleinsavak fizikai-kémiai tulajdonságai
A nukleinsavak fizikai-kémiai tulajdonságait nagy molekulatömegük és szerkezeti szerveződésük mértéke határozza meg. A nukleinsavakat a kolloid és ozmotikus tulajdonságok, az oldatok nagy viszkozitása és sűrűsége jellemzi, optikai tulajdonságok, denaturáló képesség.
A kolloid tulajdonságok minden nagy molekulatömegű vegyületre jellemzőek. Feloldódáskor a nukleinsavak megduzzadnak és viszkózus oldatokat, például kolloidokat képeznek. Hidrofilitásuk elsősorban a foszfátoktól függ. Az oldatban a nukleinsavmolekulák polianion formájúak, kifejezett savas tulajdonságokkal. Fiziológiás pH-értékeken minden nukleinsav polianion, és fehérjékből és szervetlen kationokból származó ellenionok veszik körül. A kétszálú nukleinsavak oldhatósága rosszabb, mint az egyszálúaké.
Denaturáció és renaturáció. A denaturáció azokban a makromolekulákban rejlő tulajdonság, amelyek rendelkeznek térszervezés. A denaturációt hő, expozíció okozza vegyi anyagok, amelyek megtörik a hidrogént és van der Waals kapcsolatok, stabilizálja a másodlagos és harmadlagos szerkezet nukleinsavak. Például a DNS felmelegítése a kettős hélix egyszeres láncokra oszlásához vezet, azaz „hélix-tekercs” átmenet figyelhető meg. Lassú hűtéssel a láncok újra egyesülnek a komplementaritás elve szerint. Natív DNS kettős hélix képződik. Ezt a jelenséget renaturációnak nevezik. Gyors hűtés esetén renaturáció nem következik be.
A nukleinsavak optikai aktivitásában jellemző változás kíséri denaturálódását és renaturálódását. A nukleinsavak helikális (szervezett) régiói forgatják a síkot polarizált fény, azaz optikailag aktívak, és a spirális régiók elpusztulása érvényteleníti a nukleinsavak optikai aktivitását.
Minden nukleinsavnak van maximuma optikai sűrűség körülbelül 260 nm hullámhosszon, ami megfelel a nitrogéntartalmú bázisok maximális abszorpciójának. A természetes nukleinsav felszívódásának intenzitása azonban lényegesen alacsonyabb, mint a saját nukleotidok keverékeké, amelyeket például ennek a nukleinsavnak vagy egyláncú hidrolízisével nyernek. Az ok az szerkezeti szervezet DNS és RNS, amely klasszikus hatást okoz - az optikai sűrűség csökkenése. Ezt a jelenséget hipokróm hatásnak nevezik. A legkifejezettebb azokban a nukleinsavakban, amelyek spirális szerkezetűek (például DNS) és sok GC-párt tartalmaznak (a GC-párok három hidrogénkötést tartalmaznak, ezért nehezebb felszakítani).
Nukleinsavak molekuláris hibridizációja. A nukleinsavak homológiájának vagy rokonságának meghatározására egy rendkívül fontos módszer a nukleinsavak denaturáció utáni renaturáló képességén alapul. Ezt molekuláris hibridizációnak hívják. A nukleinsavak egyszálú szakaszainak komplementer párosításán alapul.
Ez a módszer lehetővé tette a DNS elsődleges szerkezetének sajátosságainak felfedezését. Kiderült, hogy az állati DNS több (akár 100 000-szeres) ismétlődő régiót tartalmaz ugyanaz a sorrend nukleotidok. Az összes DNS 10-20%-át teszik ki. Hibridizációjuk nagyon gyorsan megy végbe. A DNS többi részét egyedi szekvenciák képviselik, amelyek nem duplikálódnak. Ezek a DNS-szakaszok nagyon lassan hibridizálnak. Egybeesésük valószínűsége az különböző organizmusok kicsi. A molekuláris hibridizáció módszerével megállapítható az egyik fajhoz tartozó szervezet DNS-ének homológiája egy másik faj DNS-ével, vagy az RNS homológiája a DNS-szelvényekkel.
Nukleinsavak és az élőlények taxonómiája
A nukleinsavak az örökletes információ anyagi hordozói, és meghatározzák az evolúció során kialakult szervezet fajspecifikusságát. A DNS nukleotid összetételének sajátosságainak tanulmányozása különböző szervezetekben lehetővé tette a taxonómiáról a külső jelek a genetikai szisztematikához. A molekuláris biológiában ezt az irányt génszisztematikának nevezik. Alapítója a kiváló szovjet biokémikus, A. N. Belozersky volt.
A DNS nukleotid-összetételének összehasonlítása különböző szervezetekben érdekes következtetésekre vezetett. Kiderült, hogy a DNS-specifitási együttható, azaz a G + C és az A + T aránya a mikroorganizmusokban nagyon változó, magasabb rendű növényeknél és állatoknál pedig meglehetősen állandó. A mikroorganizmusok ingadozásokat mutatnak az extrém GC típustól a kifejezett AT típusig. A magasabb rendű szervezetek DNS-e tartósan megtartja az AT típust. Úgy tűnhet, hogy a magasabb rendű szervezetekben a DNS specifitása elveszett. Valójában bennük ugyanolyan specifikus, mint a baktériumokban, de specifitását nem annyira a nukleotidok összetételének változékonysága, hanem a lánc mentén történő váltakozásuk sorrendje határozza meg. A DNS nukleotid-összetétele alapján érdekes következtetéseket vontak le A. N. Belozersky és tanítványai a többsejtű állatok és magasabb rendű növények eredetére vonatkozóan. AT-típusú DNS-ük áll a legközelebb a gombák DNS-éhez, így az állatok és a gombák nyilvánvalóan egy közös ősre – rendkívül primitív gombaszerű szervezetekre – vezetik vissza származásukat.
Az organizmusok rokonságáról még több információt ad a molekuláris hibridizáció módszere. Ezzel a módszerrel megállapították az emberi és majom DNS magas homológiáját. Ráadásul az emberi DNS összetételét tekintve mindössze 2-3%-ban tér el a csimpánz DNS-étől, valamivel jobban a gorilla DNS-étől, több mint 10%-ban más majmok DNS-étől, és csaknem 100%-ban a bakteriális DNS-től. A DNS elsődleges szerkezetének jellemzői a szisztematikában is felhasználhatók. Az ismétlődő szekvenciák régiói feletti homológiát (gyors hibridizáció) használják a makroszisztematikához, és az egyedi DNS-fragmensekhez (lassú hibridizáció) - a mikroszisztematikához (faj és nemzetség szintjén). A tudósok úgy vélik, hogy fokozatosan fel lehet építeni az élővilág teljes családfáját DNS segítségével.
Heteropolimereknek minősülnek?
Mi az a nukleinsav monomer?
Milyen funkcióit ismeri a nukleinsavak?
Az élőlények milyen tulajdonságait határozzák meg közvetlenül a nukleinsavak szerkezete és funkciója?
1868-ban Johann Friedrich Miescher svájci orvos és biokémikus olyan anyagot izolált az elhalt leukociták magjából, amely savanyú tulajdonságai. A tudós ezt az anyagot nukleinnek nevezte (lat. atommag– mag), tekintve, hogy csak a sejtmagokban található. Később ezeket szerves vegyületek a citoplazmában, mitokondriumokban, plasztidokban is találtak, de a nekik adott név - nukleinsavak - megmaradt.
A nukleinsavak jelentősége egy sejtben rendkívül nagy. Felépítésük sajátossága lehetővé teszi számukra, hogy ellátják az örökletes információk tárolásának, megvalósításának és továbbításának funkcióit, vagyis gyakorlatilag meghatározzák az élőlények alapvető tulajdonságait. Ezért a nukleinsavak szerkezetének tanulmányozása nagyon fontos az élő szervezetek működési elveinek megértéséhez.
Kétféle nukleinsav létezik: dezoxiribonukleinsav (DNS) és ribonukleinsav (RNS), amelyek minden sejtben megtalálhatók. Kivételt képeznek a vírusok - nem sejtes életformák, amelyek közül néhány kizárólag RNS-t, míg mások csak DNS-t tartalmaznak.
Dezoxiribonukleinsav (DNS). A 20. század közepén, amikor már bizonyított volt a DNS szerepe a tulajdonságok nemzedékről nemzedékre történő átvitelében, ezeknek a biopolimereknek a szerkezete és felépítése még teljesen tisztázatlan volt. Tudták, hogy a DNS-molekulák monomerekből – nukleotidokból – állnak, amelyek mindegyike tartalmaz egy foszforsav-maradékot, egy cukrot - dezoxiribózt és a négy nitrogénbázis egyikét - adenint (A), guanint (G), timint (T) vagy citozint (C). ); vagyis négyféle nukleotid létezik (25. ábra). De nyitva maradt az a kérdés, hogy van-e valamilyen mintázat ezeknek a monomereknek a DNS-láncban való elrendeződésében.
Az 50-es évek elején. XX század A Columbia Egyetem biokémia professzora, Erwin Chargaff a korábbi vizsgálatoknál jóval nagyobb pontossággal határozta meg a DNS összetételét. Felfedezte, hogy a tartalom négy fajta A DNS-ben lévő bázisok nem felelnek meg az 1:1:1:1 aránynak, amint azt korábban feltételeztük. A kutatót különösen megdöbbentette, hogy az adenin (A) mennyisége mindig megegyezett a timin (T), a guanin (G) mennyisége pedig mindig a citozin (C) mennyiségével. Ez nem lehetett puszta véletlen. Például az emberi DNS 30% A-t, 30% T-t, 20% G-t és 20% C-t tartalmazott. Sőt, kiderült, hogy a sejtek DNS-ének összetétele minőségileg és mennyiségileg eltérő a különböző szervezetekben, de a szervekben azonos. és ugyanannak a szervezetnek a szövetei. Ez ismét megerősítette, hogy a DNS az kémiai alaponátöröklés.
Rizs. 25. Általános képlet nukleotid (A) és négyféle DNS-nukleotid (B)
Az adenin és timin (A–T), valamint a guanin és citozin (G–C) arányának ezt a mintázatát ún. Chargaff szabályaés nyomként szolgált a DNS szerkezetéhez.
Ribonukleinsavak (RNS). Az RNS a DNS-hez hasonlóan egy biopolimer, amely négyféle monomerből – nukleotidokból – áll (28. ábra). A DNS és az RNS nukleotidjai nagyon hasonlóak, bár nem azonosak. Az RNS monomerek foszforsav-maradékot, cukor-ribózt és nitrogénbázist tartalmaznak. Ráadásul három nitrogéntartalmú bázis ugyanaz, mint a DNS-ben - adenin (A), guanin (G) és citozin (C), és a timin (T) helyett az RNS a nitrogéntartalmú uracil (U) bázist tartalmazza, amely szerkezetében hasonló. .
Az RNS nemcsak nukleotidjainak szerkezetében különbözik a DNS-től. Számos egyéb jellemző is jellemzi ezt a nukleinsavtípust.
Az RNS egyszálú molekula.
A kettős szálú RNS csak néhány RNS-tartalmú vírusban található, ahol genetikai információ tárolási funkciót lát el. A DNS és a kétszálú RNS szerkezetének hasonlósága meghatározza a funkcióbeli hasonlóságot is.

Rizs. 28. Az RNS szerkezete (R – ribóz, F – foszfátcsoport, A, U, G, C – nitrogénbázisok)
1941-ben több kutató egymástól függetlenül felfedezte, hogy a sejtek szintetizálnak nagyszámú mókus. Ez a megfigyelés arra utal, hogy az RNS fő funkciója a fehérjeszintézisben való részvétel. Később ez a hipotézis teljes mértékben beigazolódott. Sőt, kiderült, hogy egy fehérjemolekula szintéziséhez többféle RNS-re van szükség.
A szerkezettől és az elvégzett konkrét funkciótól függően Az RNS-nek három fő típusa van.
RNS átvitele (tRNS ) főként a sejt citoplazmájában található. A kis méretű, mindössze 75-90 nukleotidból álló tRNS-molekulák legfeljebb 15%-át teszik ki. teljes szám RNS a sejtben. A tRNS funkciója az aminosavak átvitele a riboszómában a fehérjeszintézis helyére.
Riboszomális RNS (rRNS ), bizonyos fehérjékhez kötődve riboszómákat képez - organellumokat, amelyek biztosítják az összes sejtfehérje szintézisét. Az rRNS-molekulák 3-5 ezer nukleotidból állnak. A sejtben található RNS összes típusa közül az rRNS teszi ki a túlnyomó többséget - körülbelül 80%.
Információ (mRNS ), vagy hírvivő RNS (mRNS ), információt ad át a fehérje szerkezetéről a DNS-ből a citoplazmában a fehérjeszintézis helyére - a riboszómába. Minden mRNS-molekula a DNS egy meghatározott szakaszának felel meg, amely egy fehérjemolekula szerkezetét kódolja. Ezért a sejt által szintetizált több ezer fehérje mindegyikéhez megvan a saját specifikus mRNS. Az mRNS mérete 300 és 30 ezer nukleotid között változik. Tól től össztömeg Az RNS egy sejt mRNS-ében 3-5%-ot tesz ki.
Minden típusú RNS szintetizálódik a DNS-en, amely egyfajta templátként szolgál e polinukleotidok létrehozásához.
Tekintse át a kérdéseket és a feladatokat
1. Mik azok a nukleinsavak? Miért kapták ezt a nevet?
2. Milyen típusú nukleinsavakat ismer?
3. Válassza ki saját kritériumait, és hasonlítsa össze a DNS- és RNS-molekulák szerkezetét. Az összehasonlító anyagot táblázat formájában mutassa be!
4. Nevezze meg a DNS funkcióit! Hogyan függenek össze a DNS szerkezete és funkciói?
5. Milyen típusú RNS található a sejtben, ahol szintetizálják őket? Sorolja fel funkcióikat.
6. Elég tudni, hogy melyik monoszacharid része a nukleotidoknak, hogy megértsük, milyen nukleinsavról beszélünk?
7. A DNS egyik szálának egy fragmentuma van következő felállás: A–G–C–G–C–C–C–T–A–. A komplementaritás elvét alkalmazva fejezze be a második láncot.
Gondol! Csináld!
1. Miért van háromféle RNS-molekula a sejtekben, de csak egyféle DNS?
2. Miben különbözik a DNS-molekula mint polimer a fehérjemolekulától?
3. Milyen típusú RNS lesz minden szervezetben azonos? Melyik RNS típus a legnagyobb variabilitással? Magyarázza el a nézőpontját.
Dolgozzon számítógéppel
Beszélni valakihez elektronikus jelentkezés. Tanulmányozza az anyagot, és oldja meg a feladatokat.
| <<< Назад
|
Előre >>> |
Előző cikk: Mekkora a fénysebesség Következő cikk: Harmonikus rezgések Az oszcillációs frekvencia fizikai képlete